

组蛋白替换是重要的染色质重塑事件之一,它在真核生物的胁迫应答中起着关键的作用。组蛋白变体取代核小体中的常规组蛋白能改变一些特定基因的转录情况,是生物响应氧化胁迫和重金属胁迫等生物或非生物胁迫的重要表达调控手段。
在过往的研究中,中科院华南植物园农业与生物技术研究中心区永祥研究员团队,发现过表达拟南芥AtOXS3(Arabidopsis OXIDATIVE STRESS 3)这种植物特异蛋白可以增强拟南芥和酵母的胁迫耐受能力。由于AtOXS3具有一个类似N-乙酰基转移酶催化结构域,并与组蛋白H4有着共同的核定位,所以该研究团队推测AtOXS3可能作为染色质重塑因子参与染色质修饰,以调控下游抗性基因表达,从而提高寄主胁迫耐受能力。
华南植物园博士研究生赖丁王等在王昌虎副研究员和区永祥研究员指导下,尝试利用模式生物裂殖酵母以证实上述推测。他们通过酵母双杂交、染色质免疫共沉淀和基因表达定量分析等手段进行了分子水平上的验证,揭示出了AtOXS3在酵母中介导胁迫抗性的相关分子机理(图1)。当酵母遇到非生物胁迫时,组蛋白H2Aα会被Tel1/Rad3激酶磷酸化并被驱逐出核小体。这时,酵母中的AtOXS3通过与组蛋白H2A的另一种变体H2A.Z及染色质重塑复合体SWR1中的Swc2亚基互作,促使相关抗药基因启动子区域上所结合的核小体中的H2Aα被H2A.Z替代。接着,转录因子Pap1被招募至这些抗药基因的启动子区域并激活其表达,从而提高细胞的胁迫耐受性。综上所述,AtOXS3作为来源于拟南芥的蛋白,它能够通过影响酵母组蛋白H2A变体的替换,特异性地激活相关抗性基因的转录。这一分子机制的阐明,不仅丰富了人们对染色质重塑的分子过程的认识,也为人们利用AtOXS3调控其它生物的胁迫应答反应提供新的思路。
相关研究成果已近期发表在国际学术期刊Genetics(《遗传学》)(2021,iyab149,IF:4.562)上。该研究获得了国家重点研发计划项目和中国科学院前沿科学重点研究项目的资助。论文链接:https://doi.org/10.1093/genetics/iyab149
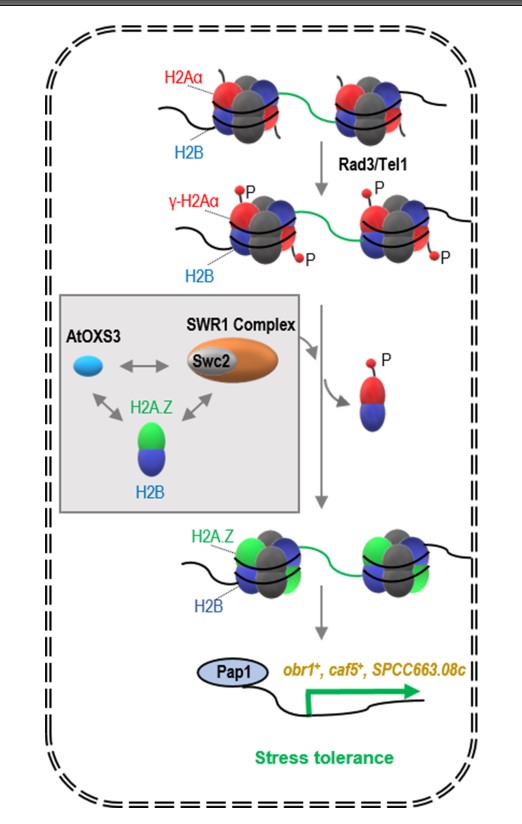
图1. AtOXS3介导胁迫耐受性的机理模型 P:磷酸化。灰色方框中显示相互作用的AtOXS3、Swc2和H2A.Z。
附件下载: